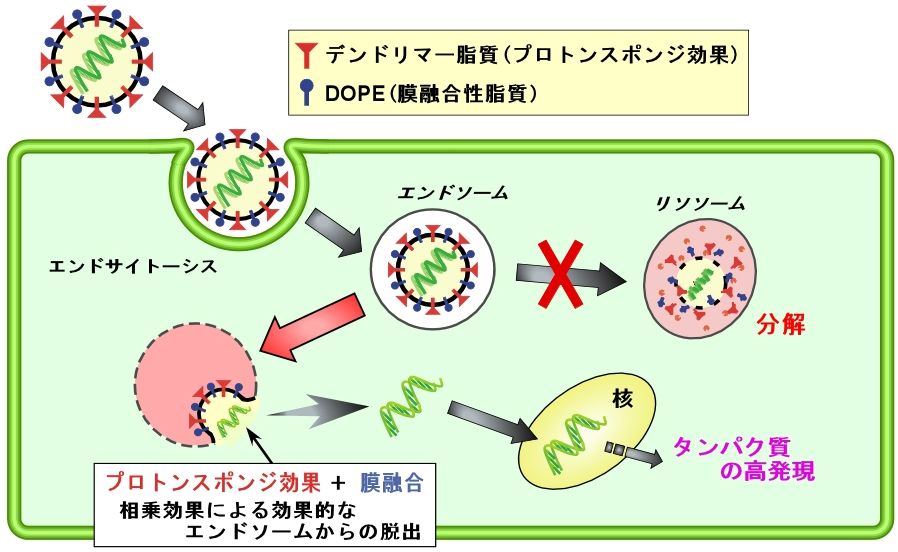
転写
-
RNAポリメラーゼの種類が多い(I, II, IIIが存在し、遺伝子認識機構と転写遺伝子がそれぞれ異なる)。
転写開始にはTATAボックスというチミン、アデニンリッチな配列が使用される。
転写終結システムは不明で、遺伝子の2000塩基対下流まで転写が進行することもある。
mRNAの修飾
-
mRNAの両末端(5'、3'両方)が修飾を受ける。
5'末端はキャップ構造が取られ、3'末端はポリアデニル化される(250個程度のアデニンをmRNA3'末端に伸長させる) イントロンをのぞくために、スプライシングを受ける。
珍しいケースとして、転写直後のmRNAとは全く別の配列になる『mRNAの編集』を受ける。
遺伝子発現調節
-
核生物の遺伝子発現調節機構は原核生物のそれより複雑で、遺伝子の上流に存在する様々な発現調節因子が関与している。
例えば、メタロチオネインというタンパク質の遺伝子は一列の上流には9個の調節因子をコードする遺伝子が知られている。
TATAボックス、GCボックス、CAATボックス、エンハンサー、サイレンサー、MREなどに、個々の遺伝子に特有な調節因子の例も多く見られる。