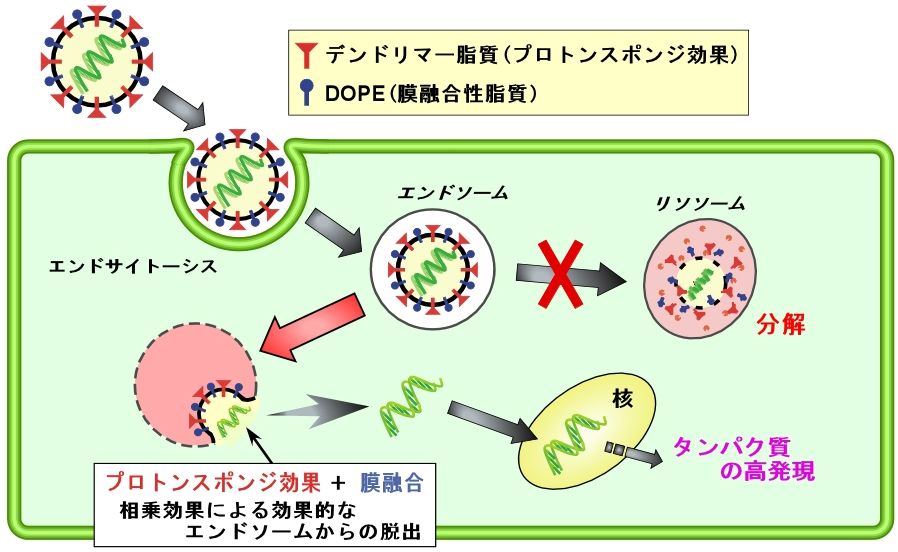
恀惓嵶嬠偱偺堚揱巕敪尰
-
堚揱巕偺敪尰偵娭偡傞懡偔偺抦尒偼恀妀惗暔偱偼側偔恀惓嵶嬠偱偁傞戝挵嬠傪儌僨儖惗暔偲偟偨幚尡偐傜摼傜傟偰偒偨丅
戝挵嬠偺堚揱巕敪尰偼婎杮揑偵埲壓偺僗僥僢僾偵暘偗傜傟傞丅
揮幨
東栿
堚揱巕惂屼乮敪尰偺挷愡乯
挷愡抜奒偼嵞傃暿偺堚揱巕敪尰偵塭嬁傪媦傏偟偨傝丄偁傞偄偼廃埻偺塰梴忦審側偳偵傛偭偰傕挷愡傪庴偗傞丅
恀惓嵶嬠偺堚揱巕敪尰條幃偼恀妀惗暔偲偼堎側傞偲偙傠偑懡偄傕偺偺丄堦斒揑側堚揱巕敪尰偲偟偰棟夝偱偒傞丅
恀惓嵶嬠偺揮幨
-
揮幨偲偼丄僎僲儉DNA偵僐乕僪偝傟傞堚揱巕杮懱偍傛傃偦偺廃曈椞堟乮嬶懱揑偵偼埲壓偵愢柧乯偑RNA億儕儊儔乕僛偵傛偭偰憡曗揑側RNA嵔 (mRNA) 偑崌惉偝傟傞夁掱偱偁傞丅昁梫側嵽椏偲偟偰偼丄
DNA埶懚惈RNA億儕儊儔乕僛
僎僲儉DNA
儕儃僰僋儗僆僠僪乮RNA暘巕:AUGC乯
埲忋偑婎杮揑側梫慺偱偁傞丅
億儕儊儔乕僛偺斀墳側偳偵偼儅僌僱僔僂儉側偳傪梫媮偡傞応崌偑偁傞偑丄偙偙偱偼妱垽偡傞丅
偙偙偱丄恀妀惗暔偲堎側傞偲偙傠偼丄RNA億儕儊儔乕僛偺峔憿偱偁傞丅
戝挵嬠偺RNA億儕儊儔乕僛偼5屄偺僒僽儐僯僢僩偐傜峔惉偝傟偰偍傝丄僒僽儐僯僢僩柤偐傜兛2兝兝乫冃峔憿乮兛僒僽儐僯僢僩丗2屄丄兝丗1屄丄兝'丗1屄丄冃丗2屄乯傪庢偭偰偄傞丅
偙傟傜偺僒僽儐僯僢僩偺栶妱偼埲壓偺傛偆偵側偭偰偄傞丅
兛僒僽儐僯僢僩丗僾儘儌乕僞乕攝楍乮屻弎乯傊偺寢崌
兝僒僽儐僯僢僩丗RNA崌惉奐巒慜偵儕儃僰僋儗僆僠僪傪愭嬱揑偵寢崌偝偣傞
兝乫僒僽儐僯僢僩丗DNA攝楍傊偺寢崌
冃僒僽儐僯僢僩丗僾儘儌乕僞乕攝楍偺擣幆
兛偼柍嶌堊偵僾儘儌乕僞乕攝楍偵寢崌偡傞偑丄冃僒僽儐僯僢僩偼偦偺攝楍傪擣幆偟偰丄敪尰偵揔摉側堚揱巕偱偁傞偐偳偆偐敾抐偡傞丅
兝偍傛傃兝'偼偦傟偧傟偑嫟栶偟偰RNA億儕儊儔乕僛妶惈傪敪婗偡傞丅
冃僒僽儐僯僢僩偼僾儘儌乕僞乕攝楍擣幆偺嵺偵昁梫偱偁傝丄揮幨偑巒傑傞偲RNA億儕儊儔乕僛偐傜棧傟偰偄偔丅
冃僒僽儐僯僢僩偑RNA億儕儊儔乕僛偵娷傑傟傞偐斲偐偱柤徧偑堎側偭偰偍傝丄埲壓偺柤慜偱帵偝傟傞丅
儂儘峺慺丗兛2兝兝乫冃峔憿丄僎僲儉DNA偺僾儘儌乕僞乕攝楍擣幆偺嵺偵峔惉偝傟傞宍忬
僐傾峺慺丗兛2兝兝乫峔憿丄揮幨偺嵟拞丄偍傛傃嵶朎撪傪梀棧偟偰偄傞忬懺偺宍忬
冃僒僽儐僯僢僩偺扙拝偼丄揮幨斀墳偵怺偔娭學偡傞丅
揮幨斀墳偼埲壓偺抜奒偵暘椶偝傟傞丅
奐巒
怢挿乮墑挿乯
廔寢
恀惓嵶嬠偺東栿
-
東栿偲偼揮幨偝傟偨mRNA偺僐僪儞乮堚揱埫崋乯偵偟偨偑偭偰丄儕儃僜乕儉偵偰傾儈僲巁偑儁僾僠僪寢崌偵傛偭偰億儕儅乕乮僞儞僷僋幙乯偵側偭偰偄偔夁掱偱偁傞丅
東栿偵昁梫側嵽椏偼丄
mRNA
傾儈僲傾僔儖tRNA乮20庬椶偺傾儈僲巁偵懳墳乯
儕儃僜乕儉
偱偁傞丅
偙偺拞偱丄摿偵恀妀惗暔偲堎側傞揰偼儕儃僜乕儉偺僒僀僘偱偁傝丄恀惓嵶嬠媦傃屆嵶嬠偱偼70S乮Svedberg扨埵乯丄恀妀惗暔偱偼80S偲側偭偰偄傞丅
傑偨丄彫僒僽儐僯僢僩丄戝僒僽儐僯僢僩偵娷傑傟傞rRNA偺挿偝傕堎側偭偰偄傞乮徻嵶偼儕儃僜乕儉傪嶲徠乯丅
東栿夁掱偼埲壓偺抜奒偵暘椶偝傟傞丅
奐巒
怢挿
廔寢